
OCR Specification focus:
‘State that TP forms carbohydrates, lipids and amino acids, and is recycled to regenerate RuBP.’
Triose phosphate (TP), produced during the Calvin cycle, is a central molecule in photosynthesis. It acts as a vital intermediate linking carbohydrate, lipid, and amino acid synthesis.
The Central Role of Triose Phosphate (TP)
Triose phosphate, also known as glyceraldehyde-3-phosphate (GALP), is the first stable, three-carbon sugar formed during the light-independent reactions of photosynthesis. It is the key output of the Calvin cycle, formed when glycerate-3-phosphate (GP) is reduced using ATP and reduced NADP generated in the light-dependent reactions. The energy and reducing power from these molecules drive the transformation of inorganic carbon into organic molecules.
Although TP can be considered the “building block” of most organic compounds in plants, its fates are carefully regulated by the plant cell’s energy requirements and the availability of carbon. Only a small proportion of TP is removed from the cycle to synthesise macromolecules, while the majority is recycled to regenerate ribulose bisphosphate (RuBP), ensuring the continuity of the Calvin cycle.
Recycling of Triose Phosphate to Regenerate RuBP
Importance of RuBP Regeneration
For carbon fixation to continue, RuBP—a five-carbon compound—must be constantly regenerated. This process maintains the supply of the molecule that reacts with carbon dioxide, catalysed by RuBisCO.
Conversion Process
Five molecules of TP (each containing three carbons) are rearranged through a series of enzymatic steps to form three molecules of RuBP (each with five carbons).
ATP from the light-dependent stage provides the energy required for phosphorylation steps that convert intermediate sugars back into RuBP.
This regeneration step consumes the majority of TP produced; only one out of every six molecules of TP typically exits the cycle.
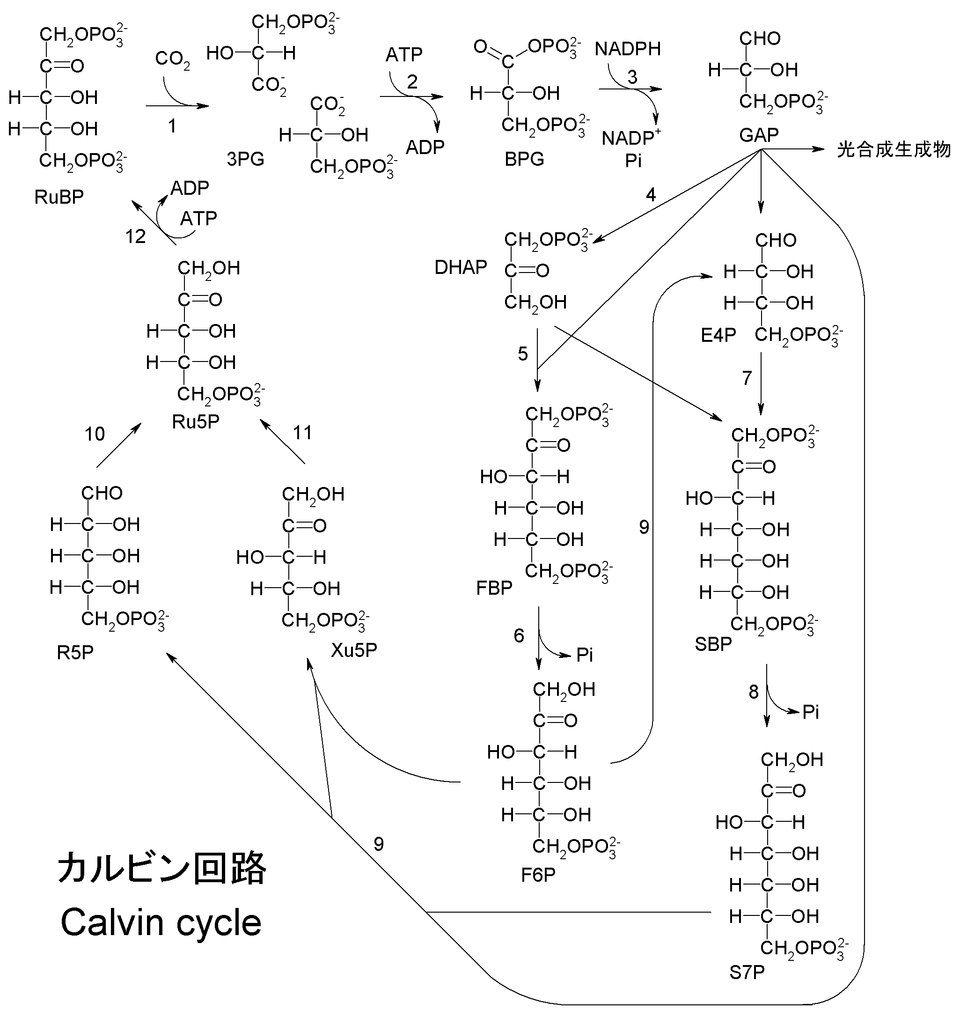
A labelled overview of the Calvin cycle, highlighting triose phosphate (G3P) formation, its recycling to RuBP, and its diversion to sucrose and starch. The visual supports the OCR requirement that TP forms carbohydrates and is recycled. Minor extra pathway detail is included but remains within A-Level scope. Source.
By maintaining this regeneration loop, the plant sustains its photosynthetic productivity. Without sufficient ATP or reduced NADP, the Calvin cycle slows, leading to reduced TP and RuBP levels.
The Biosynthetic Fates of TP
Triose phosphate serves as the universal precursor for the biosynthesis of carbohydrates, lipids, and amino acids. These pathways represent how the energy captured in photosynthesis is converted into the plant’s biochemical reserves and structural components.
1. Carbohydrate Formation
TP molecules are combined and modified to form a range of carbohydrates essential for both storage and structure.
Glucose and fructose synthesis: Two molecules of TP condense to produce hexose sugars such as glucose and fructose.
Sucrose formation: Glucose and fructose combine to form sucrose, which is translocated via the phloem to non-photosynthetic tissues.
Starch synthesis: In the chloroplast stroma, glucose units polymerise to form starch granules for long-term energy storage.
Cellulose formation: In the cell wall, glucose monomers are polymerised to cellulose microfibrils, providing mechanical strength to plant cells.
Carbohydrate: An organic compound made of carbon, hydrogen, and oxygen, often serving as an energy source or structural material in living organisms.
These carbohydrate products highlight how photosynthesis supports both the plant’s immediate metabolic needs and its longer-term energy demands.
2. Lipid Formation
Some TP is diverted into pathways leading to the synthesis of lipids, which function as energy reserves and membrane components.
TP is converted into pyruvate through glycolysis within the chloroplast or cytosol.
Pyruvate then forms acetyl-CoA, the two-carbon unit central to fatty acid synthesis.
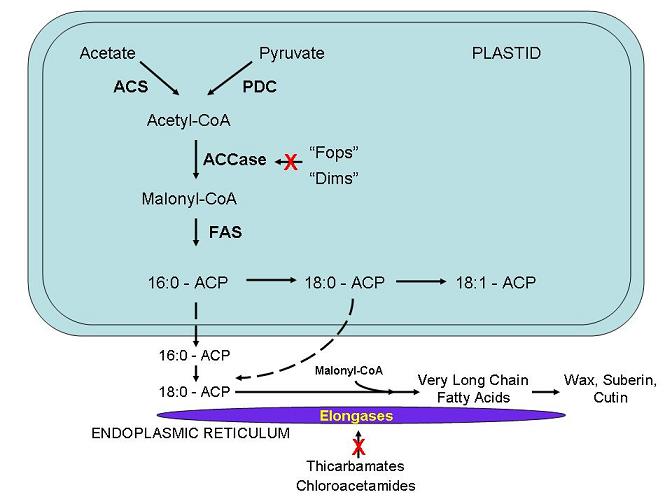
Simplified pathway of fatty-acid synthesis in plant plastids, starting from acetyl-CoA and proceeding via ACCase and fatty acid synthase (FAS) to long-chain fatty acids. This supports the OCR point that TP contributes to lipid formation (glycerol from TP; fatty acids from acetyl-CoA). The figure includes some regulatory enzyme names beyond the syllabus, noted here for context. Source.
Successive condensation reactions lengthen carbon chains, forming fatty acids, which combine with glycerol (also derived from TP) to form triglycerides and phospholipids.
Lipid: A group of hydrophobic biological molecules including fats, oils, and phospholipids, used for long-term energy storage and forming cellular membranes.
These lipid molecules contribute to the structural integrity of cell membranes and the storage of energy in seeds and other tissues.
3. Amino Acid Formation
TP also feeds into the synthesis of amino acids, the building blocks of proteins essential for enzyme function, growth, and repair.
TP combines with nitrate ions (NO₃⁻) and ammonium ions (NH₄⁺) absorbed from the soil.
Through a series of transamination and amination reactions, carbon skeletons from TP and other Calvin cycle intermediates such as 2-oxoglutarate form amino acids like glycine, serine, and alanine.
These amino acids can then be assembled into proteins, which play structural and catalytic roles in the plant.
Amino acid: An organic molecule containing an amino group (–NH₂) and a carboxyl group (–COOH), forming the monomer units of proteins.
Because these biosynthetic processes draw carbon away from the Calvin cycle, plants tightly regulate them to balance growth with energy production.
Coordination Between Anabolism and Photosynthesis
Balancing Carbon Allocation
Only around one-sixth of TP molecules leave the Calvin cycle for biosynthesis. The remaining five-sixths regenerate RuBP. This balance ensures:
Continuous carbon fixation through RuBisCO activity.
Adequate ATP and reduced NADP utilisation from the light-dependent reactions.
Controlled storage of carbohydrates, preventing osmotic imbalance in the chloroplast.
Environmental Influence
Factors such as light intensity, temperature, and CO₂ concentration influence the distribution of TP. For instance:
Under high light and adequate CO₂, excess TP promotes starch synthesis.
Under low light or CO₂ limitation, TP regeneration dominates to maintain the Calvin cycle.
During nitrogen deficiency, amino acid formation declines, causing carbon to accumulate as carbohydrate.
The Interconnected Metabolic Web
The fates of triose phosphate demonstrate how photosynthesis supports all major aspects of plant metabolism. TP acts as a metabolic hub, connecting:
Energy transfer from light to chemical form (ATP and reduced NADP).
Carbon assimilation into stable organic compounds.
Biosynthetic pathways that form the macromolecules needed for growth and survival.
Through this integration, plants efficiently convert inorganic carbon into the diverse molecules essential for life. The ability of TP to form carbohydrates, lipids, and amino acids, while simultaneously being recycled to regenerate RuBP, underpins the interdependence of photosynthetic processes described in the OCR A-Level Biology specification.
Practice Questions
Question 1 (2 marks)
State two different fates of triose phosphate (TP) produced during the Calvin cycle.
Mark Scheme:
One mark for each correctly stated fate of TP (maximum 2 marks).
Accept any two of the following:
• Used to synthesise carbohydrates (e.g. glucose, fructose, starch, or sucrose).
• Used in lipid synthesis (converted to pyruvate then acetyl-CoA).
• Used to form amino acids.
• Recycled to regenerate RuBP.
(2 marks total)
Question 2 (5 marks)
Describe how triose phosphate (TP) is used in plants to form organic molecules and how it is recycled in the Calvin cycle.
Mark Scheme:
Award marks for the following valid points, up to a maximum of 5:
• TP is a three-carbon compound formed in the Calvin cycle when glycerate-3-phosphate (GP) is reduced using ATP and reduced NADP. (1)
• Two TP molecules combine to form hexose sugars such as glucose or fructose, which can be converted into sucrose or starch. (1)
• TP can be converted into pyruvate and then acetyl-CoA, which is used to synthesise fatty acids and lipids. (1)
• TP and other Calvin cycle intermediates combine with nitrogen sources (e.g. nitrate or ammonium ions) to produce amino acids. (1)
• The majority of TP (five out of every six molecules) is recycled through a series of reactions, using ATP, to regenerate RuBP so that carbon fixation can continue. (1)
FAQ
Most TP must be recycled to regenerate RuBP, ensuring the Calvin cycle can continue to fix carbon dioxide. If too much TP were diverted to biosynthesis, RuBP levels would fall, stopping the cycle.
Plants therefore maintain a balance between using TP for growth and preserving it for RuBP regeneration. The ratio of TP used for biosynthesis versus regeneration adjusts according to light intensity, carbon dioxide concentration, and the plant’s metabolic needs.
TP moves across the chloroplast envelope via a specific triose phosphate/phosphate translocator (TPT).
For every TP molecule exported, one inorganic phosphate (Pi) is imported to maintain phosphate balance.
This exchange ensures that chloroplast ATP synthesis is not limited by phosphate depletion.
Exported TP in the cytosol is used to synthesise sucrose, which is then transported through the phloem to other tissues.
When light intensity and carbon dioxide are abundant, TP accumulates faster than it can be used for RuBP regeneration or sucrose production.
Excess TP is diverted into starch synthesis within the chloroplast.
Starch acts as a temporary carbon store and can be broken down at night when photosynthesis ceases.
This balance prevents feedback inhibition of the Calvin cycle and helps maintain steady photosynthetic rates.
TP provides the carbon skeletons that combine with inorganic nitrogen sources to produce amino acids.
Carbon compounds derived from TP, such as 2-oxoglutarate or pyruvate, act as frameworks for incorporating ammonium or nitrate ions.
Enzymes like glutamine synthetase and glutamate synthase (GOGAT) facilitate these assimilation reactions.
This process links carbon metabolism from photosynthesis directly to nitrogen metabolism required for protein synthesis.
TP is central to many cellular processes because it connects photosynthesis to respiration and biosynthesis.
In the cytosol, TP can enter glycolysis to release energy via respiration when needed.
It provides the starting material for the synthesis of sugars, fatty acids, and amino acids.
Its versatility means TP acts as a bridge between energy production, carbon fixation, and the synthesis of vital macromolecules across the plant cell.
